Molecular characterization of trypanosome cytoskeletal morphogenesis
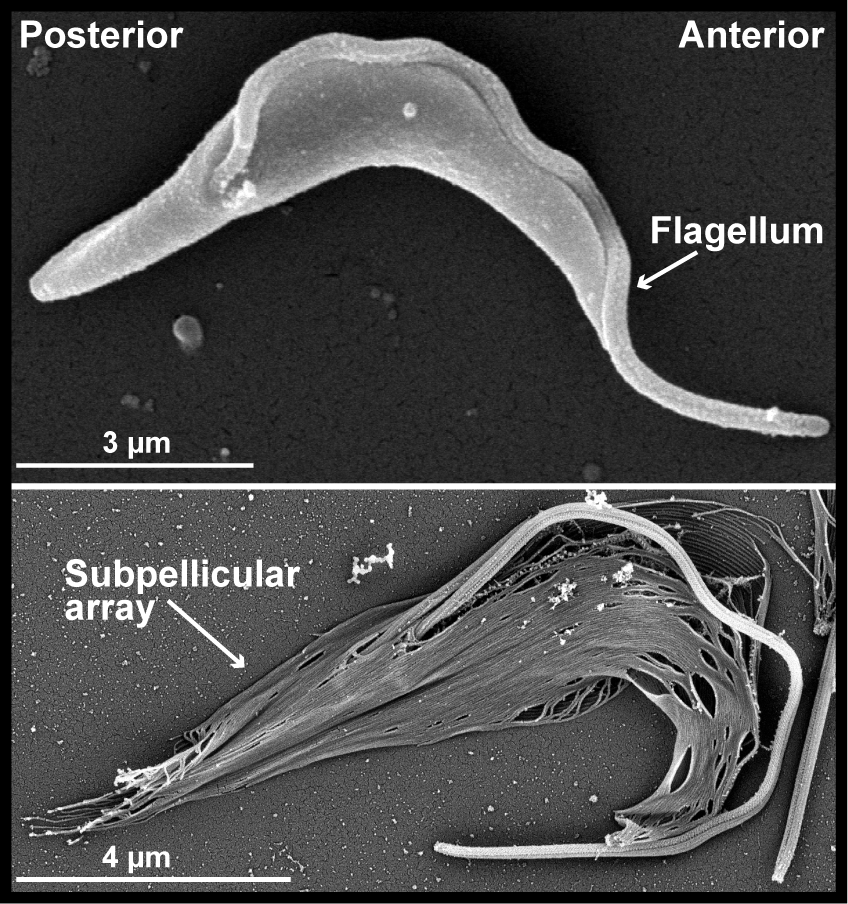
T. brucei cells are highly polarized, with distinct posterior and anterior ends. This shape is mediated by a helical array of subpellicular microtubules that underlie the plasma membrane. The flagellum is nucleated at the posterior end of the cell and is attached to the cell body, extending beyond the anterior tip of the cell. The duplication and positioning of the new flagellum during cell division is an essential morphogenic event.

T. brucei have an invagination of the plasma membrane called the flagellar pocket near the posterior of the cell that is the sole site of endo- and exo-cytosis. The flagellum is nucleated at the base of the pocket by a basal body, and the top of the compartment is tightly associated with the flagellar membrane, which limits access to the pocket lumen. A series of cytoskeletal structures are associated with the top of the flagellar pocket and are essential for formation of the seal. The flagellar pocket collar and hook complex play an important role in the construction of a new flagellar pocket, while the centrin arm appears to be essential for the formation of a new FAZ.
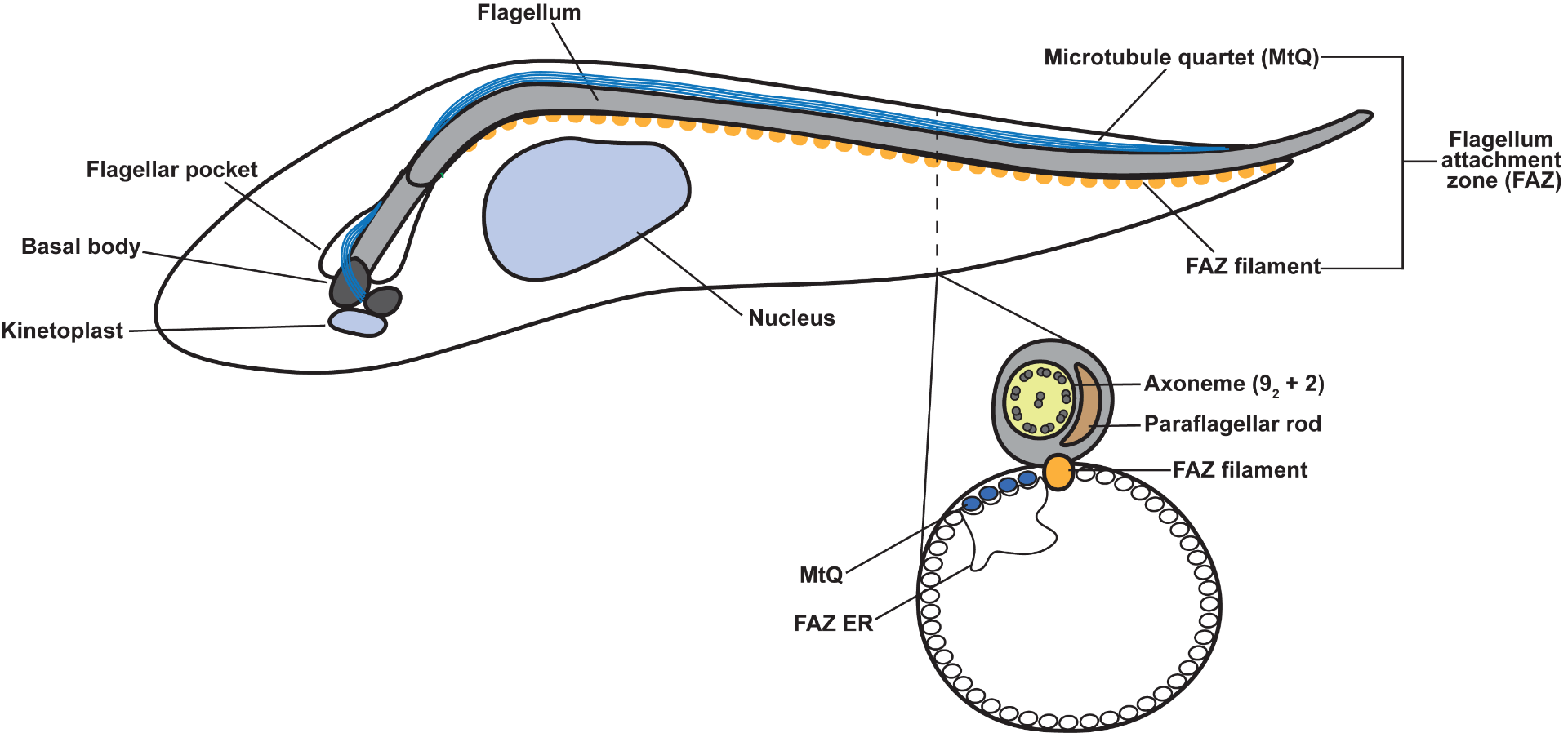
Once the flagellum emerges onto the cell surface, a series of junctional complexes known as the flagellum attachment zone (FAZ) are responsible for maintaining the positioning of the flagellum. During cell division, the tip of the new FAZ is closely associated with the old FAZ and determines the point of cleavage furrow ingression.
Our laboratory uses a variety of biochemical and cell biological techniques to 1) determine how T. brucei forms flagellar pocket-associated structures to regulate endo- and exo-cytosis and 2) discover how the subpellicular array is built and partitioned during cytokinesis.
TbPLK/TbSmee1/TOEFAZ1: molecular regulation of flagellar pocket and cytokinesis

We showed that a T. brucei homolog of polo-like kinase (TbPLK) is recruited to the basal body, then migrates to the top of the flagellar pocket, and then remains associated with the tip of the new FAZ as the structure extends. Depletion or small-molecule inhibition of TbPLK causes defects in the duplication of the flagellar pocket and FAZ, leading to cells with detached new flagella that cannot produce viable progeny.
Using a series of methods including proximity-dependent biotinylation (BioID) and phosphoproteomics, we identified a set of potential TbPLK substrates and interacting partners. These proteins include the hook complex protein TbSmee1, and the essential cytokinetic protein TOEFAZ1, which localizes to the tip of the new FAZ. Current projects in the lab focus on understanding the molecular mechanisms behind TbPLK, TbSmee1, and TOEFAZ1 function.
PAVE1 and KLIF: subpellicular array partitioning
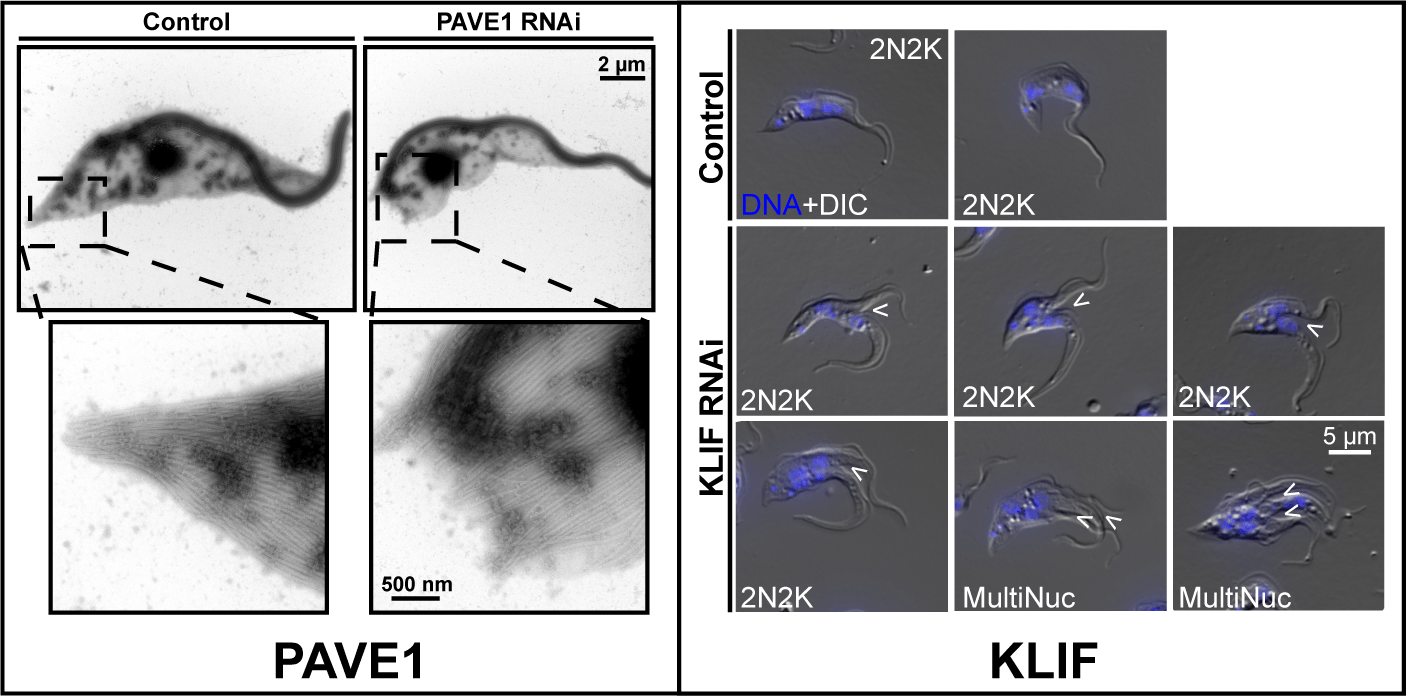
We recently performed a BioID screen to discover potential TOEFAZ1 interactors. This screen identified a set of proteins that are essential for the late stages of T. brucei cell division, when the formation of a new cell posterior occurs. We identified an essential kinesin (KLIF) that tracks along the ingressing cleavage furrow. Cells lacking KLIF are left with partially ingressed furrows and re-enter the cell cycle without ever completing this process. We also identified PAVE1, which localizes to the posterior and ventral edge of the cell. Depleting PAVE1 leads to cells that lack the tapered portion of the posterior end and are unable to correctly form the subpellicular microtubule array. This defect causes leads to cell cycle arrest. We are currently studying the function of these proteins using biophysical approaches.
Morphogenesis in other trypanosomatids
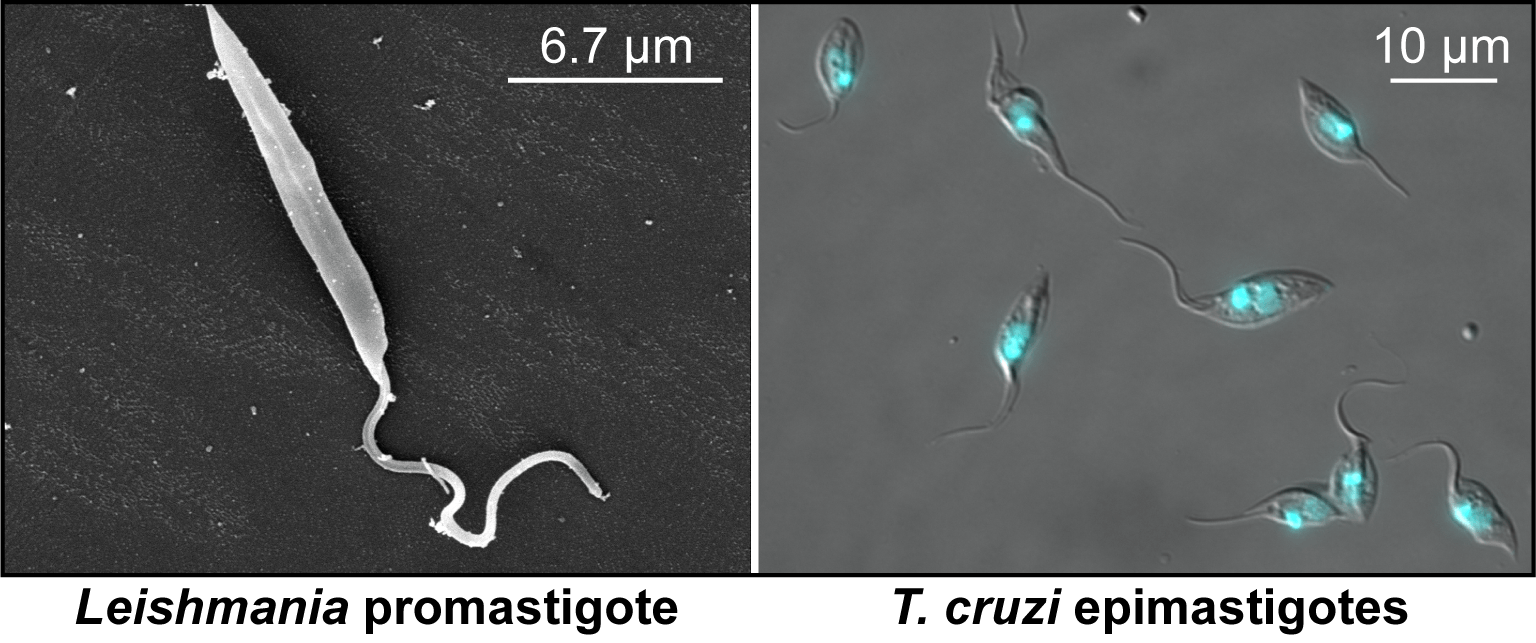
We are currently developing strategies for studying morphogenesis in Leishmania and Trypanosoma cruzi. These related parasites have retained most of the cytoskeletal proteins we have identified in T. brucei, but have unique morphologies and feature intracellular stages that are not well understood.